Bioelektriciteit: Actiepotentiaal en het model van Hodgkin en Huxley
 Spanningsafhankelijke geleidbaarheid van kaliumkanalen
Spanningsafhankelijke geleidbaarheid van kaliumkanalen
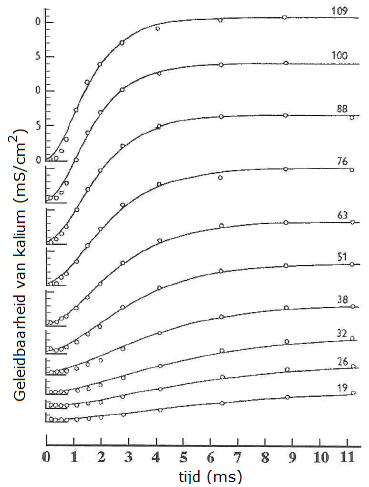
Metingen van Hodgkin en Huxley in voltage-clamp experimenten
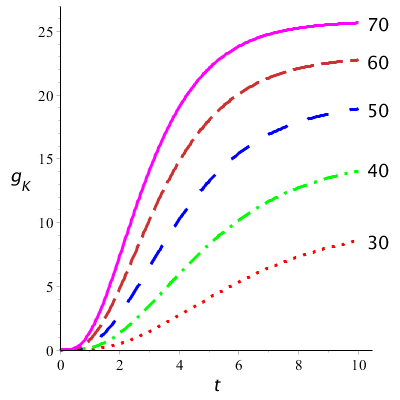
Onderstaand diagram toont de meetpunten van Hodgkin en Huxley van hun voltage-clamp experimenten waarin de geleidbaarheid van kaliumkanalen bij verschillende opgelegde spanningen in de loop van de tijd zijn uitgezet (in steeds eigen coördinatenstelsels). Bedenk dat het hier steeds een stapsgewijze verhogingen van de opgelegde spanning (rechts bij de krommen genoteerd) gaat en de reactie van het membraan hierop betreft. Voor de geleidbaarheid \(g_\mathrm{K}\) geldt: \[g_\mathrm{K}=\frac{V_m-E_\mathrm{K}}{I_\mathrm{K}}\] oftewel \[I_K=g_K(V_m-E_\mathrm{K})\] De meetgegevens bij een opgelegde spanning zijn vervolgens steeds gemodelleerd met een wiskundige formule waarvan de grafiek getekend is. De grafieken beschrijven de meetgegevens goed.
Normering van geleidbaarheid
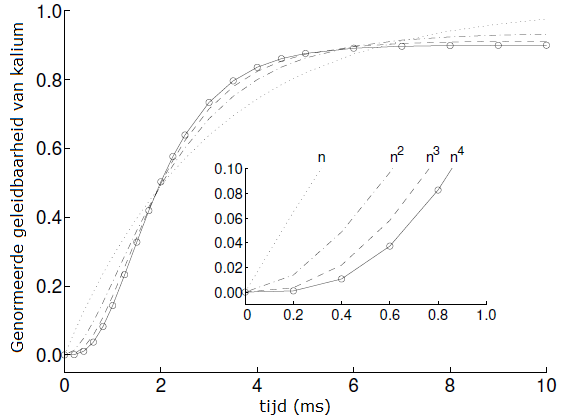
De grafiek van de geleidbaarheid \(g_\mathrm{K}\) van kaliumkanalen lijkt bij toenemende spanning steeds meer op een sigmoïde. Maar als je de grafieken normeert dan zijn ze in feite allemaal van deze vorm (zie onderstaand diagram). Normering is in feite niet veel anders dan de geleidbaarheid \(g_\mathrm{K}\) te schrijven als het product van de maximale geleidbaarheid, zeg \(\overline{g_\mathrm{K}}\) en een functie \(p_\mathrm{K}(t)\): \[g_\mathrm{K}=\overline{g_\mathrm{K}}\cdot p_\mathrm{K}(t)\] Deze functie \(p_\mathrm{K}(t)\) is de genormeerde geleidbaarheid van kaliumkanalen. Dit stelde Hodgkin en Huxley in staat om een algemene formule voor geleidbaarheid van kaliumkanalen als functie van spanning en tijd op te stellen.
Wiskundig model
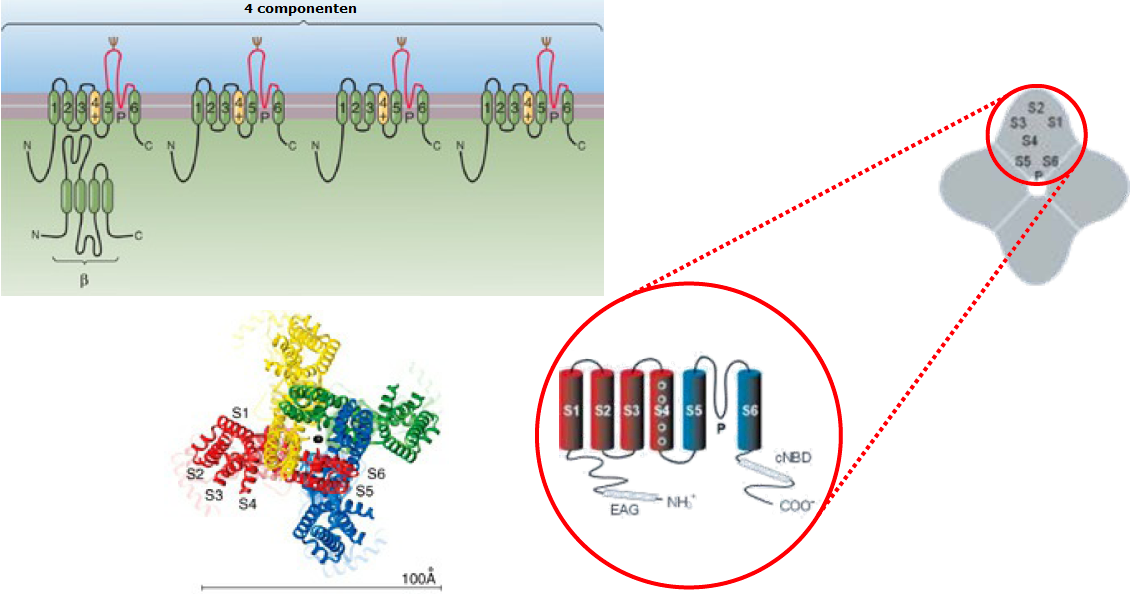
Basisidee is om eerst een wiskundig model op te stellen waarbij \(\overline{g_\mathrm{K}}\) de maximale geleidbaarheid van kaliumkanalen is wanneer alle kaliumkanalen open staan en \(p_\mathrm{K}\) de fractie van open kaliumkanalen symboliseert. Hodgkin en Huxley veronderstelden dat hiervoor vier componenten van een kaliumkanaal in doorlatende stand moeten zijn om stroom door het ionkanaal te kunnen laten stromen. De kans dat elke component afzonderlijk doorlatend is veronderstelden zij steeds door dezelfde functie \(n(t)\) beschreven te worden en ook namen zij aan dat de kansprocessen onafhankelijk zijn. Dit geeft de formule \[p_\mathrm{K}=n^4\] Naar analogie met chemische reactiekinetiek veronderstelden Hodgkin en Huxley dat er een 'evenwichtsreactie' is voor elk van de componenten van niet-doorlatend naar doorlatende toestand en vice versa met 'reactieconstanten' \(\alpha_n\) en \(\beta_n\) (deze naamkeuze wordt later wel duidelijk) in een model van elementaire reactievergelijkingen. Dus hebben we een evenwicht \[\text{niet-doorlatend}\rightleftharpoons \text{doorlatend}\] dat eigenlijk uit twee simultane processen bestaat, namelijk
voorwaarts: \(\displaystyle \text{ niet-doorlatend}\stackrel{\alpha_n}{\longrightarrow} \text{doorlatend}\) met procesconstante \(\alpha_n\)
terugwaarts: \(\displaystyle \text{ doorlatend}\stackrel{\beta_n}{\longrightarrow} \text{niet-doorlatend}\) met procesconstante \(\beta_n\)
Het veranderingsproces wordt beschreven door de volgende differentiaalvergelijking \[\frac{\dd n}{\dd t}=\alpha_n\cdot (1-n)-\beta_n\cdot n\] oftewel als \[\frac{\dd n}{\dd t} = - (\alpha_n+\beta_n)n + \alpha_n\] De differentiaalvergelijking hoort bij begrensde exponentiële groei en heeft als oplossing bij beginwaarde \(n(0)=0\) (kans op doorlatendheid gelijk aan 0): \[n(t) = n_\infty\cdot \left(1-\exp\bigl(-\frac{t}{\tau_n}\bigr)\right)\] met steady-state waarde \( n_\infty\) en tijdconstante \(\tau_h\) gegeven door \[n_\infty = \frac{\alpha_n}{\alpha_n+\beta_n}\quad\mathrm{en}\quad \tau_n=\frac{1}{\alpha_n+\beta_n}\] In onderstaand figuur staat de grafiek van de functie \(n(t)\) in het binnenkader getekend samen met de grafieken van de functies \(n^2\), \(n^3\) en \(n^4\).
Hodgkin en Huxley baseerden hun keuze van vier componenten en dus hun keuze van \(n^4\) louter en alleen op basis van een goede overeenstemming van modelresultaten met meetgegevens. Je zou kunnen zeggen dat ze het alleen maar gedaan hebben om de sigmoïdale vorm op orde te krijgen. Later is evenwel gebleken uit studies naar de structuur en de werking van het kaliumkanaal dat er inderdaad vier componenten te onderscheiden zijn. In onderstaande figuur is linksonder het bovenaanzicht van de 3-dimensionale structuur met 4 identieke componenten van een kaliumkanaal in een zoogdier te zien die samen de al dan niet gesloten holte voor kalium-ionenstroom samenstelt. Ook dit werk aan de structuur van ionkanalen door MacKinnon is in 2003 met de Nobelprijs beloond.
Spanningsafhankelijkheid
Bedenk wel dat in het model van Hodgkin en Huxley de functies \(\alpha_n\) en \(\beta_n\), en hiermee ook \(n_\infty\) en \(\tau_n\), nog afhankelijk zijn van de membraanspanning. Wat Hodgkin en Huxley dus nog restte was formules hiervoor op te stellen op basis van hun meetgegevens. Zij stelden (in eenheden van \(\mathrm{ms}^{-1}\) en \(\mathrm{mV}\)) voor: \[\alpha_n = \frac{0.01(10-v)}{\exp\left(\frac{10-v}{10}\right)-1}\quad\mathrm{en}\quad \beta_n=0.125\exp\left(\frac{-v}{80}\right)\] waarbij \(v\) het verschil tussen de membraanspanning en de rustmembraanspanning \(V_r=-70\;\mathrm{V}\) is. Als je dus de spanningsafhankelijkheid niet ten opzichte van de rustmembraanspanning wilt bestuderen maar ten opzicht van de membraanpotentiaal dan moet je voor \(v\) invullen \(V-V_r=V+70\).
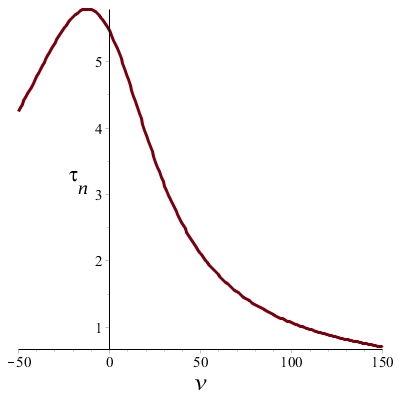
Wanneer we bovenstaande formules toepassen dan kunnen we de tijdconstante \(\tau_n\) en de steady-state waarde \(n_\infty\) uitzetten tegen de spanning \(v\) en zo de spanningsafhankelijkheid in grafiekvorm zien:
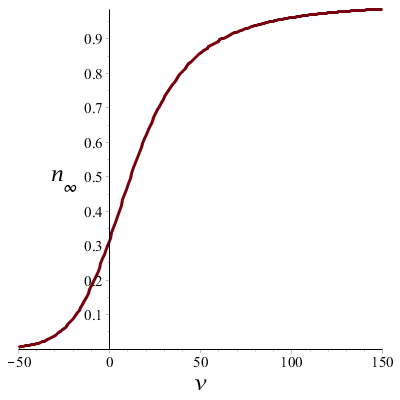
De onderste grafiek laat zien dat bij depolarisatie (positieve \(v\)) de kaliumgeleidbaarheid toeneemt als de spanning groter wordt en het kaliumkanaal neigt naar een doorlatende toestand. Bij sterk negatieve potentiaal neigt het kaliumkanaal tot sluiting.
De bovenste grafiek laat zien dat ingeval van depolarisatie de tijdconstante \(\tau_n\) afneemt naarmate de spanning groter wordt; met andere woorden, naarmate de depolarisatie groter is zal de cel sneller hierop 'reageren'. De tijdconstante is het grootst rond de rustmembraanpotentiaal en daalt aan weerszijden.
Hieronder zijn grafieken getekend van de modelkrommen van kaliumgeleidbaarheid uitgezet tegen tijd voor verschillende waarden van \(v\). Hierbij is gebruik gemaakt van de volgende waarde voor de maximale geleidbaarheid: \(g_\mathrm{K} =36\;\mathrm{mS\,cm}^{-2}\). Ze zien er uit zoals in het eerste diagram van metingen gedaan door Hodgkin en Huxley.